ABSTRACT
Hepatocellular carcinoma (HCC) is a highly lethal cancer in which epigenetic dysregulation plays a key role in tumor development and progression. Among epigenetic mechanisms, histone methylation regulates chromatin structure and gene transcription and influences multiple aspects of HCC biology, including tumor proliferation, metastasis, immune modulation, cancer stemness, and therapeutic resistance. In this review, we summarize recent advances in understanding the roles of histone methyltransferases in HCC, focusing on their involvement in tumor proliferation, metastasis, immune regulation, drug resistance, and cancer stem cell maintenance. We also discuss their clinical relevance as potential diagnostic and prognostic biomarkers and highlight emerging therapeutic strategies targeting histone methylation pathways. Collectively, these findings suggest that histone methyltransferases represent promising targets for developing novel epigenetic-based diagnostic and therapeutic approaches for HCC.
-
KEYWORDS: Hepatocellular carcinoma; Histone methylation; Epigenetic regulation; Therapeutic target; Histone methyltransferase
INTRODUCTION
Hepatocellular carcinoma (HCC) is one of the most common and lethal malignancies worldwide, characterized by complex genetic and epigenetic alterations that drive tumor initiation and progression [
1,
2]. Among these mechanisms, histone methylation has emerged as a key epigenetic modification that regulates chromatin structure and gene transcription, thereby influencing multiple biological processes involved in cancer development [
3,
4]. Accumulating evidence indicates that histone methyltransferases and related regulators control critical aspects of HCC biology, including tumor proliferation, metastasis, immune modulation, therapeutic resistance, and cancer stem cell maintenance [
5]. Recent studies have demonstrated that dysregulation of histone methylation enzymes such as enhancer of zeste homolog 2 (EZH2), SET domain bifurcated histone lysine methyltransferase 1 (SETDB1), SET and MYND domain–containing protein (SMYD) family members, protein arginine methyltransferases (PRMTs), and lysine methyltransferases (KMT) family proteins contributes to hepatocarcinogenesis by regulating oncogenic transcriptional programs, metabolic pathways, and tumor microenvironment signaling [
6,
7]. These epigenetic regulators interact with non-coding RNAs, immune signaling pathways, and viral factors to further promote tumor progression and treatment resistance [
8,
9]. In addition, the expression of histone methylation regulators is frequently associated with tumor aggressiveness, poor prognosis, and altered responses to targeted therapies and immunotherapies [
10].
Thus, in this review, we provide a comprehensive overview of recent advances in histone methyltransferases and regulators in HCC, integrating their diverse functions into a unified framework of tumor progression and therapeutic response. We further discuss their clinical relevance as diagnostic and prognostic biomarkers, along with emerging therapeutic strategies targeting histone methylation pathways.
PROLIFERATION
Histone methyltransferases (HMTs) are central to the epigenetic control of gene expression during HCC progression, shaping chromatin accessibility and transcriptional programs. These HMTs coordinate multiple oncogenic processes, including metabolic adaptation and DNA damage responses, that support tumor cell growth and survival.
Mixed-lineage leukemia 1 (MLL1)–menin complex maintains oncogenic transcription in HCC by sustaining H3K4me3 deposition and chromatin accessibility at target genes, and inhibition of the menin–MLL1 interaction reduces H3K4me3 levels and gene expression, impairing HCC cell survival [
11]. KMT5C, an H4K20 trimethyltransferase, promotes HCC progression by interacting with RAD51 recombinase (RAD51) to enhance homologous recombination–mediated DNA damage repair and tumor cell survival; its inhibition sensitizes HCC cells to poly (ADP-ribose) polymerase (PARP) inhibitors [
12]. Conversely, loss of KMT2C (MLL3) promotes HCC development by reducing H3K4 methylation at the cyclin-dependent kinase inhibitor 2A (CDKN2A) locus, suppressing p16/Ink4a and p19/Arf expression, while restoration of KMT2C reactivates CDKN2A transcription and induces tumor cell apoptosis [
13].
Several regulators interact with histone methylation machinery to influence HCC progression. Splicing factor 3b subunit 4 (SF3B4) promotes tumor growth by regulating gene expression and alternative splicing, and its interaction with SETD5 links RNA splicing to histone methylation–associated transcriptional regulation in HCC, while SETD5, frequently upregulated in HCC, enhances cell proliferation, invasion, and glycolysis through pyruvate kinase, muscle (PKM) regulation, and its depletion suppresses tumor growth and inflammatory signaling [
14,
15]. Ubiquitin specific peptidase 27 (USP27) promotes hepatocarcinogenesis by deubiquitinating and stabilizing SETD3, thereby enhancing tumor cell proliferation and tumorigenesis, with high USP27–SETD3 expression correlating with poor patient prognosis [
16]. In contrast, spectrin beta, non-erythrocytic 1 (SPTBN1) functions as a tumor suppressor by promoting SETD7-mediated methylation of yes-associated protein 1 (YAP1), leading to YAP degradation and suppression of oncogenic signaling, whereas loss of SPTBN1 stabilizes YAP and promotes tumor progression [
17].
Euchromatic histone lysine methyltransferase 2 (EHMT2; G9a) is frequently overexpressed in MYC proto-oncogene, bHLH transcription factor (MYC) -driven HCC and mediates H3K9 methylation–dependent gene repression, contributing to hepatocarcinogenesis by interacting with and stabilizing c-Myc, which promotes tumor growth and invasion and correlates with poor patient prognosis [
18]. In addition, G9a suppresses apoptosis by repressing the pro-apoptotic gene BCL2-like 14 (BCL2L14; Bcl-G) through inhibition of p53-mediated transcription during the DNA damage response, allowing damaged hepatocytes to evade apoptosis and facilitating tumor initiation; accordingly, pharmacological inhibition of G9a reduces H3K9me2 levels, restores apoptotic signaling, and suppresses liver tumor initiation and growth [
19,
20].
Members of the SMYD family of methyltransferases play important roles in HCC progression through diverse epigenetic and metabolic mechanisms. SMYD2 promotes tumor growth by methylating c-Myc, increasing its stability and activating the c-Myc–glutaminase 1 (GLS1) axis, thereby enhancing glutamine metabolism and contributing to sorafenib resistance in HCC cells [
21]. SMYD3, frequently upregulated in HCC, promotes proliferation and invasion by interacting with the nucleosome remodeling and deacetylase complex (NuRD) (MTA1/2) complex, increasing H4K20me3 and reducing histone acetylation at tumor-suppressive gene promoters, and also activates oncogenic signaling by catalyzing H3K4me3 at the sphingosine-1-phosphate receptor 1 (S1PR1) promoter, enhancing S1PR1 expression and correlating with poor patient prognosis [
22,
23]. Beyond transcriptional regulation, SMYD5 promotes tumor growth by catalyzing ribosomal protein L40 (RPL40) K22 trimethylation, which enhances translation elongation and global protein synthesis and is associated with poor prognosis in HCC patients [
24]. In addition, SMYD4 acts as an oncogenic lysine methyltransferase by methylating PRMT5, enhancing its activity and promoting H3R2 and H4R3 symmetric dimethylation, thereby activating oncogenic gene expression and driving HCC proliferation and metastasis [
25].
EZH2 promotes HCC progression mainly through H3K27me3-mediated epigenetic gene silencing, enhancing tumor cell proliferation and survival and regulating oncogenic transcriptional programs such as the EZH2–minichromosome maintenance (MCM)–Human telomerase reverse transcriptase (hTERT) axis, which suppresses senescence and supports continuous tumor growth; disruption of this pathway reduces hTERT expression and increases sensitivity to sorafenib [
26,
27]. EZH2 also modulates major signaling pathways including wnt family member (WNT) /β-catenin, creating therapeutic vulnerability to WNTinib in β-catenin– activated HCC [
28]. In addition, polycomb repressive complex 2 (PRC2)-dependent gene repression by EZH2 limits the efficacy of DNA methyltransferase inhibitors, whereas combined inhibition of EZH2 and DNA methyltransferase (DNMT) reactivates silenced genes and enhances anti-tumor effects [
29].
PRMT family methyltransferases regulate HCC progression through distinct histone arginine methylation mechanisms. PRMT1 promotes HCC progression by catalyzing H4R3me2a, which epigenetically activates SRY-box transcription factor 18 (SOX18) transcription, leading to increased proliferation, migration, and invasion and correlating with poor patient prognosis [
30]. Coactivator-associated arginine methyltransferase 1 (CARM1), frequently overexpressed in HCC, catalyzes H3R17me2 methylation to activate fermitin family member 1 (FERMT1) transcription and drive tumor proliferation and metastasis, while pharmacological inhibition of CARM1 suppresses malignant behaviors in HCC cells [
31]. In contrast, PRMT5 suppresses hepatocarcinogenesis by catalyzing H4R3me2s to repress P21 transcription; loss of PRMT5 increases P21 expression, causing pathological polyploidization and liver cirrhosis that promote HCC development [
32].
Non-coding RNAs regulate EZH2-mediated epigenetic pathways during HCC progression. The lncRNA SNHG9, highly expressed in HCC and liver cancer stem cells, promotes cancer stem cell (CSC) self-renewal and tumorigenesis by recruiting EZH2 to increase H3K27me3 at the phosphatase and tensin homolog (PTEN) promoter, leading to suppression of the tumor suppressor PTEN, whereas inhibition of small nucleolar RNA host gene 9 (SNHG9) restores PTEN expression and suppresses tumor growth [
33]. In contrast, miR-15a functions as a tumor suppressor, where P53-induced miR-15a suppresses O-linked N-acetylglucosamine (O-GlcNAc) transferase (OGT), reducing O-GlcNAc–mediated stabilization of EZH2 and thereby inhibiting HCC cell proliferation, migration, invasion, and tumor progression through the P53–miR-15a–OGT–EZH2 axis [
34].
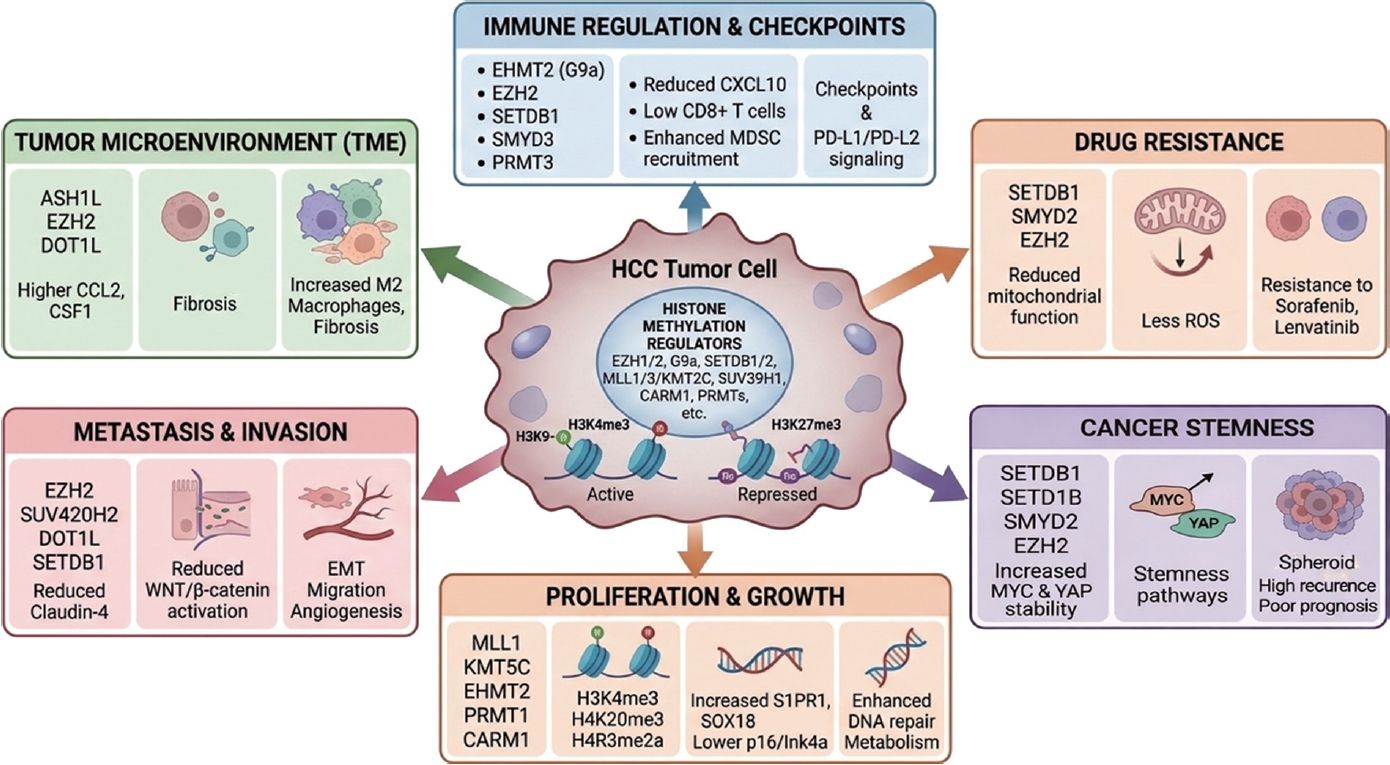
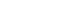
Overall, histone methylation–driven regulatory networks integrate transcriptional, metabolic, and DNA repair pathways to sustain tumor proliferation. Targeting these coordinated epigenetic programs may provide effective strategies for limiting tumor growth in HCC (
Table 1).
METASTASIS AND TUMOR MICROENVIRONMENT (TME)
Histone methylation regulators modulate key processes underlying HCC dissemination, including epithelial–mesenchymal transition (EMT), migration, and invasion. In addition to intrinsic tumor cell changes, these epigenetic mechanisms influence interactions between cancer cells and the surrounding microenvironment.
EZH2 promotes HCC metastasis by increasing H3K27me3-mediated repression of the claudin-4 gene, which activates Wnt/β-catenin signaling and lymphoid enhancer-binding factor 1 (LEF-1)–dependent EMT, leading to increased expression of metastasis markers such as matrix metallopeptidase 9 (MMP-9) and vimentin; inhibition of EZH2 restores claudin-4 expression, alters EMT-related gene expression including E-cadherin, N-cadherin, and vimentin, and suppresses metastatic potential [
35,
36]. Oxidative stress–induced suppressor of variegation 4-20 homolog 2 (SUV420H2) upregulation increases H4K20me3, which promotes EMT, migration, and invasion of HCC cells and correlates with poor prognosis, while inhibition of H4K20me3 formation suppresses tumor progression [
37]. Disruptor of telomeric silencing 1-like (DOT1L) also promotes HCC metastasis by inducing EMT-associated transcriptional programs, although macrophage-mediated nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling counteracts the anti-metastatic effect of DOT1L inhibition in vivo, and combined targeting of DOT1L and macrophage/NF-κB pathways enhances therapeutic efficacy [
38].
In addition, several lncRNA-mediated epigenetic pathways regulate HCC metastasis. ESET (SETDB1) suppresses the tumor-suppressive lncRNA GMDS-AS1 through H3K9 methylation, leading to increased expression of the oncogenic target proteasome subunit beta type-1 (PSMB1) and promoting proliferation and metastasis [
39], whereas LINC01348, a tumor-suppressive lncRNA, inhibits HCC growth and metastasis by regulating EZH2 alternative splicing through splicing factor 3b subunit 3 (SF3B3) and modulating the c-Jun N-terminal kinase (JNK)/Jun proto-oncogene, AP-1 transcription factor subunit (c-JUN)/snail family transcriptional repressor 1 (Snail) signaling pathway, while C-terminal truncated hepatitis B virus X protein (Ct-HBx) downregulates LINC01348 via c-Jun signaling to promote metastasis [
40]. Furthermore, the hypoxia-induced lncRNA SZT2-AS1 enhances HCC growth, metastasis, and angiogenesis by recruiting hypoxia-inducible factor (HIF)-1α/β and SMYD2 to hypoxia response elements, increasing H3K4me3 and H3K36me3 and activating transcription of HIF target genes [
41].
Histone methylation–associated pathways influence the composition and function of immune cells within the tumor microenvironment. Consequently, histone methylation– related mechanisms contribute to immune modulation and immune evasion in HCC. Glutamate signaling promotes HCC progression by downregulating EZH2, which decreases H3K27me3 at the C–C motif chemokine ligand 2 (CCL2) promoter and increases CCL2 expression, thereby enhancing recruitment of tumor-associated macrophages (TAMs) and promoting tumor progression [
42]. EZH2 inhibition also alters immune cell composition, increasing recruitment of myeloid-derived suppressor cells (MDSCs) through chemokine induction and suppressing CD8⁺ T cell–mediated anti-tumor immunity, while combined EZH2 inhibition and myeloid-derived suppressor cell (MDSC) depletion restores anti-tumor responses [
43]. In addition, absent, small, or homeotic discs 1-like (ASH1L) promotes fibrosis-associated HCC by catalyzing H3K4me3 to increase CCL2 and colony-stimulating factor 1 (CSF1) expression, which recruits and polarizes M2-like macrophages and establishes an immunosuppressive tumor microenvironment; inhibition or depletion of ASH1L reduces fibrosis, tumor growth, and immune suppression in HCC [
44].
Together, histone methylation–dependent mechanisms link tumor-intrinsic programs with microenvironmental remodeling, coordinating metastatic progression and immune suppression. These dual functions highlight their importance as integrative drivers of HCC progression (
Table 2).
IMMUNE RESPONSE
Histone methyltransferases and regulators critically influence anti-tumor immunity by shaping immune cell infiltration, checkpoint signaling, and tumor–immune interactions. These epigenetic pathways act as key regulators of immune responsiveness within the HCC microenvironment.
G9a suppresses anti-tumor immunity by catalyzing H3K9me2 at the C–X–C motif chemokine ligand 10 (CXCL10) promoter, reducing chemokine expression and limiting CD8⁺ T cell recruitment, whereas G9a inhibition restores CXCL10 expression and enhances responsiveness to anti– programmed cell death protein 1 (anti–PD-1) immunotherapy [
45]. SETDB1 overexpression suppresses immune cell infiltration and immune pathway activity, potentially reducing the effectiveness of immune checkpoint therapy in HCC [
46]. SMYD3 promotes tumor progression by inducing PDL2 expression through H3K4me3 under extracellular matrix stiffness, which suppresses xCT-mediated ferroptosis and contributes to tumor growth and sorafenib resistance, while targeting the SMYD3–PD-L2 axis enhances ferroptosis and therapeutic responses [
47]. Wolf–hirschhorn syndrome candidate 1 (WHSC1; NSD2) is upregulated in HCC and catalyzes H3K36me2, regulating transcription through epigenetic crosstalk with H3K27me3 and DNA methylation, and is associated with DNA repair, cell cycle regulation, immune cell infiltration, and poor prognosis [
48]. Alterations in SETD2 may influence immunotherapy response, as mutations were associated with exceptional responses to ipilimumab and nivolumab, suggesting SETD2 as a potential predictive biomarker [
49]. EZH2 is frequently overexpressed in HCC and correlates with poor prognosis, with stage-dependent DNA methylation regulating its expression and influencing immune responses and metabolic pathways [
50]; EZH2 also regulates immune cell infiltration and CD8⁺ T cell function via the macrophage migration inhibitory factor (MIF)–CD74 signaling pathway, linking EZH2 activity to prognosis and immunotherapy response, while pharmacological targeting of EZH2 (e.g., AZD6738) enhances CD8⁺ T cell proliferation and anti-tumor immunity [
51,
52]. In addition, PRMT3 promotes immune evasion by methylating pyruvate dehydrogenase kinase 1 (PDHK1), increasing lactate production that induces PD-L1 expression through H3K18 lactylation, thereby suppressing CD8⁺ T cell infiltration and promoting tumor growth via the PRMT3– PDHK1–lactate–PD-L1 axis [
53]. Finally, the SNHG6 or metastasis-associated lung adenocarcinoma transcript 1 (MALAT1)/miR-101-3p/EZH2 regulatory axis contributes to EZH2 upregulation by suppressing the EZH2-targeting miRNA miR-101-3p, representing a non-coding RNA-mediated pathway that promotes HCC progression and immune microenvironment modulation [
54].
Collectively, histone methyltransferases and regulators are key modulators of the tumor immune microenvironment in HCC, governing immune cell infiltration, immune checkpoint pathways, and mechanisms of immune evasion (
Table 3). Aberrant activity of enzymes such as G9a, SETDB1, EZH2, and PRMT3 alters anti-tumor immune responses and impacts the efficacy of immunotherapy, underscoring histone methylation pathways as promising targets for enhancing therapeutic outcomes in HCC.
DRUG RESISTANCE AND STEMNESS
Epigenetic regulators play important roles in drug resistance and cancer stemness in HCC. Histone methyltransferases and related pathways influence tumor cell survival, stemness maintenance, and therapeutic sensitivity. These epigenetic mechanisms contribute to resistance to targeted therapies and chemotherapy in HCC.
SETDB1-high tumors exhibit enhanced stemness, dedifferentiation, reduced immune infiltration, and resistance to immunotherapy and conventional treatments, indicating a role in cancer stem cell maintenance and immune evasion, and SETDB1 also modulates sorafenib sensitivity by regulating mitochondrial function and dynamin related protein 1 (DRP1)-mediated mitochondrial dynamics, suggesting its potential as a predictive marker for sorafenib response in HCC [
55,
56]. Additionally, SETD1B is upregulated in HCC and promotes liver cancer stem cell (LCSC) stemness through the MYC-associated zinc finger protein (MAZ)–SETD1B–CD24 axis, enhancing tumor sphere formation, adhesion, and tumor growth, while targeting SETD1B with triptolide suppresses LCSC properties and inhibits HCC progression [
57]. And, SMYD2 promotes sorafenib resistance and cancer stemness by activating bone morphogenetic protein 4 (BMP4) transcription through H3K4me1 and H3K36me2, thereby driving the SMYD2–BMP4–SMAD1/5/8–ID3 signaling axis associated with poor prognosis [
58]. EZH2 contributes to therapeutic resistance in HCC through multiple epigenetic mechanisms. EZH2-mediated H3K27me3 represses ferroptosis-related genes, including acyl-CoA synthetase long chain family member 1 (ACSL1) in lenvatinib resistance and transferrin receptor 2 (TFR2) in sorafenib resistance, and also suppresses cysteine–methionine metabolism genes such as betaine–homocysteine s-methyltransferase (BHMT) and cysteine dioxygenase type 1 (CDO1), thereby regulating lipid metabolism and ferroptosis susceptibility; inhibition of EZH2 restores ferroptosis, increases lipid peroxidation, and sensitizes HCC cells to targeted therapies [
59-
61]. In addition, EZH2 promotes drug resistance through signaling pathways, including infigratinib resistance via Erb-B receptor tyrosine kinase (ErbB) 2/ErbB3 signaling and fibroblast growth factor receptor 4 (FGFR4) inhibitor resistance through NF-κB–mediated EZH2 accumulation and YAP activation, whereas combined targeting of EZH2 with ErbB or FGFR4 inhibitors enhances apoptosis and suppresses HCC progression [
62,
63].
Non-coding RNAs regulate histone methyltransferase– mediated oncogenic pathways and drug resistance in HCC. EZH2 promotes HCC progression and chemoresistance by suppressing miR-381 through H3K27me3, leading to SETDB1 upregulation and activation of the AKT signaling pathway via K64 trimethylation, which enhances tumor proliferation, migration, tumorigenesis, and chemotherapy resistance [
64]. In contrast, several microRNAs act as tumor suppressors: miR-20a directly targets EZH1, and pharmacological inhibition of EZH1/EZH2 with UNC1999 suppresses tumor growth and enhances sorafenib sensitivity, highlighting the miR-20a–EZH1 axis as a therapeutic target [
65]. Similarly, miR-138 directly targets EZH2, and its overexpression suppresses EZH2-mediated EMT, thereby increasing cisplatin sensitivity and inhibiting tumor progression in HCC [
66].
Thus, histone methyltransferase and regulators contribute to drug resistance and cancer stemness in HCC by regulating tumor cell survival, metabolic adaptation, and stemness-related signaling pathways. Targeting key regulators such as SETDB1, SMYD2, and EZH2, as well as their non-coding RNA– mediated networks, may represent promising strategies to overcome therapeutic resistance and improve treatment outcomes in HCC (
Table 4).
PROGNOSIS
Aberrant expression of histone methylation regulators is closely associated with disease progression and clinical outcomes in hepatocellular carcinoma. These epigenetic factors reflect tumor aggressiveness and underlying biological heterogeneity. SETD1A promotes cancer stem cell stemness by catalyzing H3K4me3 and activating oncogenic enhancers and super-enhancers, simultaneously inducing oncogenes and repressing tumor suppressor genes, with elevated SETD1A expression correlating with poor prognosis and HCC relapse [
67]. SETDB1 is frequently upregulated in HCC and correlates with tumor size, microvascular invasion, and advanced pTNM stage, suggesting its role in tumor progression and its potential as a prognostic indicator [
68]. SUV420H1 (KMT5B), an H4K20 histone methyltransferase, is also overexpressed in HCC and promotes cell cycle progression, invasion, and tumor growth, while its inhibition suppresses tumor proliferation and metastasis [
69]. In addition, EZH2 is frequently overexpressed in HCC and associated with poor survival, higher recurrence rates, advanced tumor stage, and elevated AFP levels, while also regulating tumor-associated macrophage infiltration, DNA recombination, and cell cycle–related pathways, and its knockdown significantly reduces HCC cell viability [
6,
70]. Similarly, EZH1 overexpression is linked to poor prognosis and drug resistance, promoting tumor progression through autophagy-related pathways such as autophagy related 7 (ATG7) activation [
71]. Thus, histone methyltransferases serve as key determinants of tumor behavior and clinical prognosis, supporting their utility as biomarkers for risk stratification and disease outcome prediction in HCC (
Table 5).
OTHERS
Histone methyltransferases regulate cell proliferation, DNA damage responses, and key signaling pathways involved in tumor progression. Suppressor of variegation 3-9 homolog 1 (SUV39H1) is upregulated by hepatitis B virus (HBV) infection and promotes HCC cell proliferation and migration by enhancing mitochondrial oxidative phosphorylation (OXPHOS), and elevated serum SUV39H1 levels may serve as a diagnostic biomarker for HBV-associated HCC [
72]. In contrast, KMT2D functions as a tumor suppressor, where the tet methylcytosine dioxygenase 2 (TET2)–KMT2D axis promotes transcription of KMT2D and AT-rich interaction domain 1A (ARID1A) to maintain tumor-suppressive epigenetic signaling, while loss of KMT2D enhances tumor growth and vitamin C–mediated activation of this pathway suppresses HCC progression [
73]. HBV DNA integration frequently occurs at the KMT2B locus, enhancing hepatic cell proliferation through H3K4me3-mediated activation of cell-cycle genes and inducing replication stress and DNA damage, thereby contributing to hepatocarcinogenesis in HBV-related HCC [
74]. In addition, EZH2 promotes tumor progression by regulating circadian rhythm–related genes such as circadian locomotor output cycles kaput (CLOCK) and cryptochrome circadian regulator 1 (CRY1), and its knockdown suppresses proliferation and induces DNA damage in HCC cells [
75]. EZH2 also regulates therapy resistance, as it suppresses miR-138-5p via H3K27me3, activating the HIF-1α signaling pathway, whereas miR-138-5p overexpression or EZH2 knockdown enhances radiosensitivity and inhibits migration, invasion, and EMT in HCC cells [
76]. Thus, epigenetic regulators contribute to HCC progression through diverse mechanisms, including viral infection, metabolic regulation, and transcriptional control, influencing tumor development and therapeutic response (
Table 5).
CONCLUSION
Histone methyltransferases play central and multifaceted roles in HCC by regulating key biological processes, including tumor proliferation, metastasis, immune modulation, and therapeutic resistance. Representative enzymes such as EZH2, SETDB1, SMYD family members, PRMTs, and KMT proteins contribute to the control of oncogenic transcriptional programs, metabolic reprogramming, and the maintenance of cancer stem cell properties. In addition, these enzymes influence the tumor microenvironment by modulating immune responses and cellular interactions within the tumor niche. Accumulating evidence indicates that dysregulation of histone methyltransferases is closely associated with tumor initiation, progression, metastasis, recurrence, and unfavorable clinical outcomes in HCC. Given their critical functional roles and clinical relevance, histone methyltransferases have emerged as promising targets for translational and clinical applications. Several of these enzymes hold potential as diag-nostic and prognostic biomarkers, enabling more precise patient stratification. Furthermore, pharmacological inhibition of histone methyltransferases has shown encouraging results in restoring tumor suppressor pathways, reprogramming aberrant epigenetic states, and enhancing the efficacy of existing therapeutic strategies, including targeted therapy and immunotherapy. Therefore, continued investigation into histone methylation–mediated regulatory mechanisms is expected to advance our understanding of HCC pathogenesis and to support the development of novel diagnostic tools, prognostic indicators, and epigenetic therapeutic approaches (
Fig. 1).
NOTES
-
ACKNOWLEDGEMENTS
Not applicable.
-
FUND
This work was supported by a National Research Foundation of Korea (NRF) grant funded by the Ministry of Science, ICT and Future Planning (RS-2018-NR056534, RS-2023-00225239, RS-2024-00336620, RS-2025-25443867, RS-2026-25520117, RS-2026-25515390, JDC0032611) and by the KRIBB Research Initiative Program (KQM0042611, KGM1062612). The funders had no role in the study design, data collection or analysis, decision to publish, or manuscript preparation.
-
ETHICS STATEMENT
Consent for publication is not required, as this submission does not include any images or information that could identify any individual.
-
CONFLICTS OF INTEREST
No potential conflict of interest relevant to this article was reported.
Figure 1.Graphical summary of histone methylation–mediated regulation in HCC. Histone methylation regulators control multiple aspects of HCC biology, including tumor proliferation, metastasis, tumor microenvironment remodeling, immune regulation, drug resistance, and cancer stemness. These epigenetic pathways ultimately influence disease progression and clinical outcomes and represent promising targets for diagnostic and therapeutic strategies in HCC.
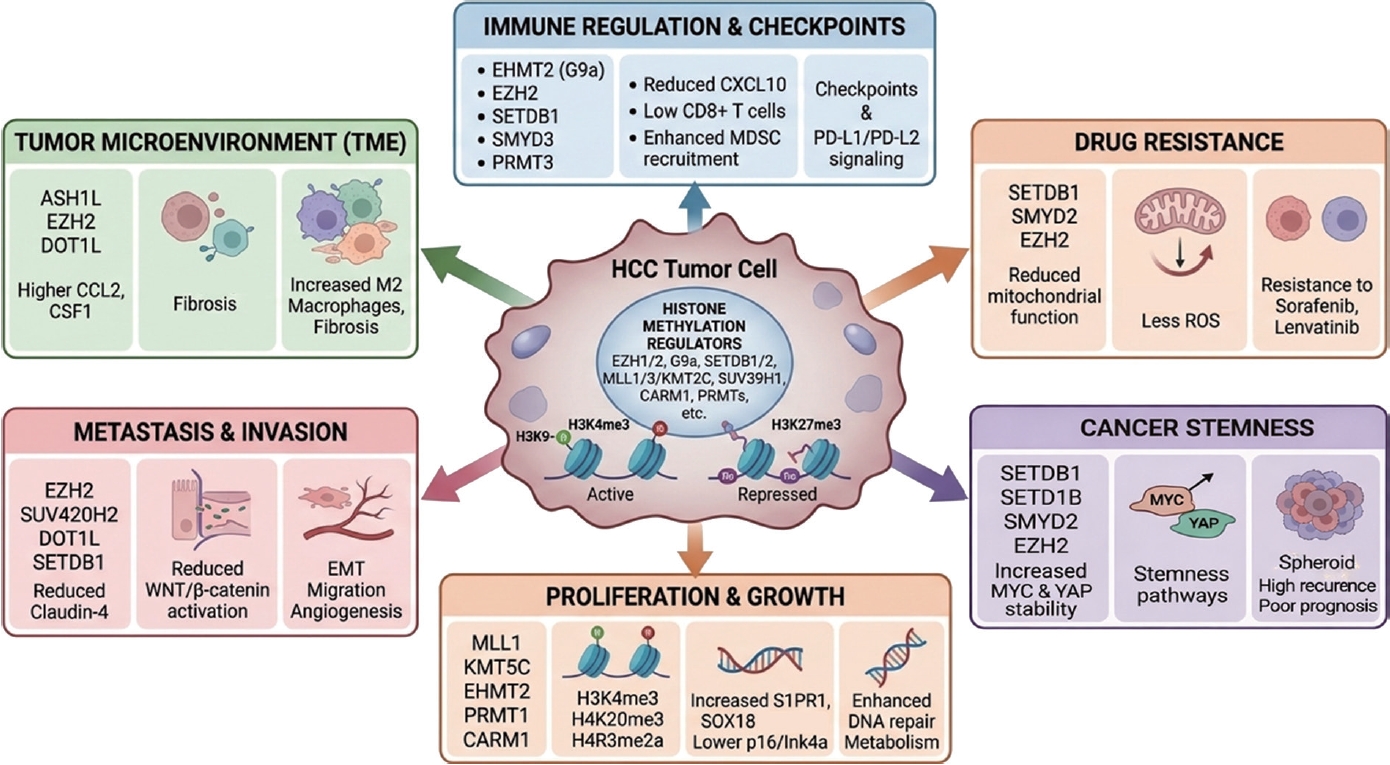
Table 1.Key histone methyltransferases involved in HCC proliferation
Table 1.
|
Gene |
Mechanism |
Reference |
|
MLL1 |
Maintains oncogenic transcription via H3K4me3 and chromatin accessibility |
[11] |
|
KMT5C |
Enhances DNA damage repair via RAD51 interaction, promoting cell survival |
[12] |
|
KMT2C |
Loss suppresses CDKN2A, promoting tumorigenesis |
[13] |
|
SETD5 |
Promotes proliferation and glycolysis via PKM regulation |
[14,15] |
|
EHMT2 |
Induces H3K9 methylation, stabilizes c-Myc, and suppresses apoptosis (Bcl-G/p53 axis) |
[18-20] |
|
SMYD2 |
Stabilizes c-Myc and enhances glutamine metabolism and drug resistance |
[21] |
|
SMYD3 |
Activates oncogenic transcription via H3K4me3 and NuRD complex interaction |
[22,23] |
|
SMYD5 |
Promotes translation and protein synthesis via RPL40 methylation |
[24] |
|
SMYD4 |
Activates PRMT5 to enhance oncogenic gene expression |
[25] |
|
EZH2 |
Silences tumor suppressors via H3K27me3, regulates WNT signaling, and cooperates with DNMT |
[26-29] |
|
PRMT1 |
Activates SOX18 via H4R3me2a, promoting proliferation and invasion |
[30] |
|
CARM1 |
Activates FERMT1 via H3R17me2, enhancing tumor progression |
[31] |
|
PRMT5 |
Represses P21 via H4R3me2s (loss promotes tumorigenesis) |
[32] |
Table 2.Histone methyltransferases governing metastasis and the tumor microenvironment in HCC
Table 2.
|
Gene |
Mechanism |
Reference |
|
EZH2 |
Represses claudin-4 via H3K27me3, activating Wnt/β-catenin–driven EMT and metastasis; modulates immune cell recruitment (TAMs, MDSCs) and suppresses CD8+ T cell responses |
[35,36,42,43] |
|
Suv420h2 |
Induces H4K20me3 under oxidative stress, promoting EMT, migration, and invasion |
[37] |
|
DOT1L |
Activates EMT-associated transcription; efficacy enhanced by combined inhibition with macrophage/NF-κB signaling |
[38] |
|
SETDB1 (ESET) |
Suppresses tumor-suppressive lncRNA (GMDS-AS1) via H3K9 methylation, promoting metastasis |
[39] |
|
ASH1L |
Induces CCL2/CSF1 via H3K4me3, promoting M2 macrophage polarization and immunosuppressive TME |
[44] |
Table 3.Histone methylation–mediated regulation of immune response in HCC
Table 3.
|
Gene |
Mechanism |
Reference |
|
EHMT2 |
Represses CXCL10 via H3K9me2, reducing CD8+ T cell recruitment and anti-tumor immunity (inhibition enhances anti-PD-1 response) |
[45] |
|
SETDB1 |
Suppresses immune cell infiltration and immune pathway activity, reducing immunotherapy efficacy |
[46] |
|
SMYD3 |
Induces PD-L2 via H3K4me3, suppressing ferroptosis and promoting immune evasion and drug resistance |
[47] |
|
WHSC1 (NSD2) |
Catalyzes H3K36me2, regulating immune infiltration, DNA repair, and tumor progression |
[48] |
|
SETD2 |
Mutation associated with enhanced response to immune checkpoint inhibitors (predictive biomarker) |
[49] |
|
EZH2 |
Regulates immune cell infiltration, CD8+ T cell function (MIF–CD74 axis), and immunotherapy response; controlled by DNA methylation |
[50-52] |
|
PRMT3 |
Promotes PD-L1 expression via lactate production (H3K18 lactylation), suppressing CD8+ T cell infiltration |
[53] |
Table 4.Roles of histone methyltransferases in drug resistance and cancer stemness in HCC
Table 4.
|
Gene |
Mechanism |
Reference |
|
SETDB1 |
Promotes stemness, immune evasion, and sorafenib resistance via mitochondrial regulation and CSC maintenance |
[55,56] |
|
SETD1B |
Enhances liver cancer stemness via MAZ–SETD1B–CD24 axis, promoting tumor growth |
[57] |
|
SMYD2 |
Activates BMP4 signaling (SMAD1/5/8–ID3), promoting stemness and sorafenib resistance |
[58] |
|
EZH2 |
Suppresses ferroptosis (ACSL1, TFR2) and metabolic genes, and activates oncogenic pathways (ErbB, FGFR4, NF-κB/YAP), promoting drug resistance |
[59-63] |
|
EZH2 / SETDB1 axis |
EZH2 suppresses miR-381, leading to SETDB1 upregulation and AKT activation, enhancing chemoresistance |
[64] |
|
EZH1/EZH2 |
Targeted by miR-20a and inhibitor (UNC1999), restoring drug sensitivity |
[65] |
|
EZH2 |
Targeted by miR-138, suppressing EMT and increasing cisplatin sensitivity |
[66] |
Table 5.Histone methyltransferases associated with prognosis and additional oncogenic mechanisms in HCC
Table 5.
|
Gene |
Mechanism |
Reference |
|
SETD1A |
Activates oncogenic enhancers (H3K4me3) and CSC stemness, correlating with poor prognosis and relapse |
[67] |
|
SETDB1 |
Promotes tumor progression and correlates with tumor size, invasion, and advanced stage |
[68] |
|
SUV420H1 (KMT5B) |
Enhances cell cycle progression, invasion, and tumor growth |
[69] |
|
EZH2 |
Associated with poor survival, recurrence, and advanced stage; regulates TAM infiltration, DNA repair, circadian genes, and radiosensitivity |
[6,70,75,76] |
|
EZH1 |
Promotes tumor progression and drug resistance via autophagy (ATG7) |
[71] |
|
SUV39H1 |
Upregulated by HBV; enhances OXPHOS, proliferation, and migration; potential diagnostic biomarker |
[72] |
|
KMT2D |
Tumor suppressor via TET2–KMT2D axis; loss promotes tumor growth |
[73] |
|
KMT2B |
Activated by HBV integration; induces H3K4me3-mediated cell cycle activation and DNA damage |
[74] |
REFERENCES
- 1. Fernández-Barrena MG, Arechederra M, Colyn L, Berasain C, Avila MA. Epigenetics in hepatocellular carcinoma development and therapy: The tip of the iceberg. JHEP Rep 2020;2:100167.
- 2. Toh TB, Lim JJ, Chow EK. Epigenetics of hepatocellular carcinoma. Clin Transl Med 2019;8:13.
- 3. Marei HE. Epigenetic regulators in cancer therapy and progression. NPJ Precis Oncol 2025;9:206.
- 4. Zhao Z, Shilatifard A. Epigenetic modifications of histones in cancer. Genome Biol 2019;20:245.
- 5. Bueloni B, Garcia Fernandez de Barrena M, Avila MA, Bayo J, Mazzolini G. Epigenetic mechanisms involved in hepatocellular carcinoma development and progression. eGastroenterology 2025;3:e100186.
- 6. Bae AN, Jung SJ, Lee JH, Lee H, Park SG. Clinical value of EZH2 in hepatocellular carcinoma and its potential for target therapy. Medicina (Kaunas) 2022;58:155.
- 7. Song J, Yang P, Chen C, et al. Targeting epigenetic regulators as a promising avenue to overcome cancer therapy resistance. Signal Transduct Target Ther 2025;10:219.
- 8. Ramalingam PS, Zhang L, Hussain MS, et al. Non-coding RNAs as key regulators in hepatitis B virus-related hepatocellular carcinoma. Front Immunol 2025;16:1602252.
- 9. Saadi W, Fatmi A, Pallardó FV, García-Giménez JL, Mena-Molla S. Long non-coding RNAs as epigenetic regulators of immune checkpoints in cancer immunity. Cancers (Basel) 2022;15:184.
- 10. Zhu Y, Liu K, Zhu H. Targeting epigenetic methylation: emerging diagnosis and therapeutic strategies in cancer. Exp Hematol Oncol 2026;15:27.
- 11. Dzama-Karels M, Sokolowski M, Kuhlers P, Brinkman JA, Morris JP, Raab JR. Menin-MLL1 complex cooperates with NF-Y to promote hepatocellular carcinoma survival. Cell Rep 2025;44:116619.
- 12. Tong Y, Wang F, Li S, et al. Histone methyltransferase KMT5C drives liver cancer progression and directs therapeutic response to PARP inhibitors. Hepatology 2024;80:38-54.
- 13. Zhu C, Soto-Feliciano YM, Morris JP, et al. MLL3 regulates the CDKN2A tumor suppressor locus in liver cancer. Elife 2023;12:e80854.
- 14. Huang H, Fang Y, Li Z, et al. SF3B4 regulates proliferation and apoptosis in hepatocellular carcinoma via alternative splicing and interaction with TRIM28 and SETD5. J Transl Med 2025;23:441.
- 15. Park M, Moon B, Kim JH, et al. Downregulation of SETD5 suppresses the tumorigenicity of hepatocellular carcinoma cells. Mol Cells 2022;45:550-563.
- 16. Zou T, Wang Y, Dong L, et al. Stabilization of SETD3 by deubiquitinase USP27 enhances cell proliferation and hepatocellular carcinoma progression. Cell Mol Life Sci 2022;79:70.
- 17. Chen S, Wu H, Wang Z, et al. Loss of SPTBN1 suppresses autophagy via SETD7-mediated YAP methylation in hepatocellular carcinoma initiation and development. Cell Mol Gastroenterol Hepatol 2022;13:949-973.e7.
- 18. Thng DKH, Hooi L, Toh CCM, et al. Histone-lysine N-methyltransferase EHMT2 (G9a) inhibition mitigates tumorigenicity in Myc-driven liver cancer. Mol Oncol 2023;17:2275-2294.
- 19. Nakatsuka T, Tateishi K, Kato H, et al. Inhibition of histone methyltransferase G9a attenuates liver cancer initiation by sensitizing DNA-damaged hepatocytes to p53-induced apoptosis. Cell Death Dis 2021;12:99.
- 20. Nishigaya Y, Takase S, Sumiya T, et al. Discovery of novel substrate-competitive lysine methyltransferase G9a inhibitors as anticancer agents. J Med Chem 2023;66:4059-4085.
- 21. Xu K, Ding J, Zhou L, et al. SMYD2 promotes hepatocellular carcinoma progression by reprogramming glutamine metabolism via c-Myc/GLS1 axis. Cells 2022;12:25.
- 22. Yang Y, Qiu R, Zhao S, et al. SMYD3 associates with the NuRD (MTA1/2) complex to regulate transcription and promote proliferation and invasiveness in hepatocellular carcinoma cells. BMC Biol 2022;20:294.
- 23. Zhang H, Zheng Z, Zhang R, et al. SMYD3 promotes hepatocellular carcinoma progression by methylating S1PR1 promoters. Cell Death Dis 2021;12:731.
- 24. Miao B, Ge L, He C, et al. SMYD5 is a ribosomal methyltransferase that catalyzes RPL40 lysine methylation to enhance translation output and promote hepatocellular carcinoma. Cell Res 2024;34:648-660.
- 25. Zhou Z, Chen Z, Zhou Q, et al. SMYD4 monomethylates PRMT5 and forms a positive feedback loop to promote hepatocellular carcinoma progression. Cancer Sci 2024;115:1587-1601.
- 26. Bai J, Zhao B, Ma Y, Wang L, Feng P, Hua Y. Antisense-mediated exon skipping targeting EZH2 suppresses tumor growth in a xenograft mouse model of hepatocellular carcinoma. Mol Ther 2025;33:1485-1501.
- 27. Shen Z, Wang Y, Gao J, et al. The EZH2/MCM Complex/hTERT axis facilitates hepatocellular carcinoma progression by inhibiting cellular senescence. Mech Ageing Dev 2025;224:112040.
- 28. Hung MH, Wang XW. Targeting WNT/beta-Catenin via modulating EZH2 function: A new chapter in the treatment of beta-Catenin mutant hepatocellular carcinoma? Cancer Res 2023;83:3498-3500.
- 29. Zhang L, Li HT, Shereda R, et al. DNMT and EZH2 inhibitors synergize to activate therapeutic targets in hepatocellular carcinoma. Cancer Lett 2022;548:215899.
- 30. Ling J, Wang S, Yi C, et al. PRMT1-mediated modification of H4R3me2a promotes liver cancer progression by enhancing the transcriptional activity of SOX18. Hepatol Commun 2025;9:e0647.
- 31. Lu J, Wu H, Zhan P, et al. PSMD14-mediated deubiquitination of CARM1 facilitates the proliferation and metastasis of hepatocellular carcinoma by inducing the transcriptional activation of FERMT1. Cell Death Dis 2025;16:141.
- 32. Wang J, Huang X, Zheng D, Li Q, Mei M, Bao S. PRMT5 determines the pattern of polyploidization and prevents liver from cirrhosis and carcinogenesis. J Genet Genomics 2023;50:87-98.
- 33. Yang S, Ruan X, Hu B, Tu J, Cai H. lncRNA SNHG9 enhances liver cancer stem cell self-renewal and tumorigenicity by negatively regulating PTEN expression via recruiting EZH2. Cell Tissue Res 2023;394:441-453.
- 34. You Z, Peng D, Cao Y, et al. P53 suppresses the progression of hepatocellular carcinoma via miR-15a by decreasing OGT expression and EZH2 stabilization. J Cell Mol Med 2021;25:9168-9182.
- 35. Özel M, Kilic E, Baskol M, Akalın H, Baskol G. The effect of EZH2 inhibition through DZNep on epithelial-mesenchymal transition mechanism. Cell Reprogram 2021;23:139-148.
- 36. Verma S, Yadav M, Verma S, Srivastava AK, Mugale MN. EZH2-mediated hypermethylation of H3K27me3 downregulates claudin-4 and upregulates the Wnt/beta-catenin signaling pathway in hepatocellular carcinoma metastasis. Biochim Biophys Acta Mol Cell Res 2026;1873:120076.
- 37. Phoyen S, Sanpavat A, Ma-On C, et al. H4K20me3 upregulated by reactive oxygen species is associated with tumor progression and poor prognosis in patients with hepatocellular carcinoma. Heliyon 2023;9:e22589.
- 38. Yang YB, Wu CY, Wang XY, et al. Targeting inflammatory macrophages rebuilds therapeutic efficacy of DOT1L inhibition in hepatocellular carcinoma. Mol Ther 2023;31:105-118.
- 39. Huang J, Zhong T, Li G, Wang S, Qin R. Epigenetic inhibition of lncRNA GMDS-AS1 by methyltransferase ESET promoted cell viability and metastasis of hepatocellular carcinoma. Clin Transl Oncol 2023;25:1793-1804.
- 40. Lin YH, Wu MH, Liu YC, Lyu PC, Yeh CT, Lin KH. LINC01348 suppresses hepatocellular carcinoma metastasis through inhibition of SF3B3-mediated EZH2 pre-mRNA splicing. Oncogene 2021;40:4675-4685.
- 41. Liu R, Guo Y, Wang L, et al. A novel hypoxia-induced lncRNA, SZT2-AS1, boosts HCC progression by mediating HIF heterodimerization and histone trimethylation under a hypoxic microenvironment. Cell Death Differ 2025;32:714-729.
- 42. Chen J, Sun HW, Wang RZ, et al. Glutamate promotes CCL2 expression to recruit tumor-associated macrophages by restraining EZH2-mediated histone methylation in hepatocellular carcinoma. Oncoimmunology 2025;14:2497172.
- 43. Qiang N, Ao J, Nakamura M, et al. Alteration of the tumor microenvironment by pharmacological inhibition of EZH2 in hepatocellular carcinoma. Int Immunopharmacol 2023;118:110068.
- 44. Du Y, Wu S, Xi S, et al. ASH1L in hepatoma cells and hepatic stellate cells promotes fibrosis-associated hepatocellular carcinoma by modulating tumor-associated macrophages. Adv Sci (Weinh) 2024;11:e2404756.
- 45. Chai X, Chen J, Zhao Y, et al. G9a epigenetically suppresses CXCL10 expression and inhibits anti-tumor immunity in hepatocellular carcinoma. Int J Cancer 2026;158:1960-1974.
- 46. Yin CQ, Song CQ. Tumor intrinsic immunogenicity suppressor SETDB1 worsens the prognosis of patients with hepatocellular carcinoma. Cells 2024;13:2102.
- 47. Wang S, Yuan X, Yang Z, et al. Matrix stiffness-dependent PD-L2 deficiency improves SMYD3/xCT-mediated ferroptosis and the efficacy of anti-PD-1 in HCC. J Adv Res 2025;73:265-282.
- 48. Yan J, Zhang MY, Lin J, et al. WHSC1 is involved in DNA damage, cellular senescence and immune response in hepatocellular carcinoma progression. J Cell Mol Med 2023;27:1436-1441.
- 49. Reiter FP, Rau M, Kunzmann V, et al. Profound tumor response to combined CTLA-4 and PD-1 inhibition in systemic fourth line therapy observed in a patient with hepatocellular carcinoma harboring SETD2 and LRP1B mutations. Z Gastroenterol 2023;61:71-75.
- 50. Chien YC, Wu GW, Wu JY, Liu LC, Hsieh YH, Yu YL. Stage-dependent EZH2 methylation correlates with immune polarization, metabolic suppression, and unfavorable outcomes in hepatocellular carcinoma. Int J Med Sci 2025;22:4201-4213.
- 51. Li RP, Wu GZ, Fang XD, et al. Single-cell transcriptional analysis reveals the mechanism of AZD6738 in HCC immunotherapy via EZH2 targeting. Drug Des Devel Ther 2025;19:2897-2920.
- 52. Zhou Y, Xu Y, Ye M, et al. Integrated single-cell RNA-seq analysis reveals that EZH2 regulates the MIF-CD74 axis to modulate T cell activation and exhaustion in hepatocellular carcinoma. J Transl Med 2025;23:1040.
- 53. Ding CH, Yan FZ, Xu BN, et al. PRMT3 drives PD-L1-mediated immune escape through activating PDHK1-regulated glycolysis in hepatocellular carcinoma. Cell Death Dis 2025;16:158.
- 54. Chen Z, Lin X, Wan Z, et al. High expression of EZH2 mediated by ncRNAs correlates with poor prognosis and tumor immune infiltration of hepatocellular carcinoma. Genes (Basel) 2022;13:876.
- 55. Fan M, Wu J, Wu Y, et al. SETDB1 decline promotes the resistance to sorafenib via DRP1 phosphorylation-mediated mitochondrial dysfunction in HepG2 cells. Biochem Biophys Res Commun 2025;768:151934.
- 56. Padelli M, Desterke C, Devocelle A, Uzan G, Lemoine A, Giron-Michel J. The epigenetic regulator SETDB1 as a key component of cancer stem cells and drug resistance in primary liver cancer. Cell Oncol (Dordr) 2026;49:18.
- 57. Gao Y, Zhou W, Gao Y, et al. Histone methyltransferase SETD1B maintains cancer stem cell niche by regulating the crosstalk between CD24 and surface adhesion molecules in hepatocellular carcinoma. Int J Biol Sci 2025;21:4798-4815.
- 58. Wang S, Wu W, Shi Z, et al. SMYD2 epigenetically activates BMP4/SMAD1/5/8/ID3 axis to enhance cancer stem cell properties and drive sorafenib resistance in hepatocellular carcinoma. Neoplasia 2025;67:101203.
- 59. Lai Y, Han X, Xie B, et al. EZH2 suppresses ferroptosis in hepatocellular carcinoma and reduces sorafenib sensitivity through epigenetic regulation of TFR2. Cancer Sci 2024;115:2220-2234.
- 60. Lee J, You C, Kwon G, et al. Integration of epigenomic and transcriptomic profiling uncovers EZH2 target genes linked to cysteine metabolism in hepatocellular carcinoma. Cell Death Dis 2024;15:801.
- 61. Zhang Y, Lin Y, Cai H, Zhou T. EZH2 confers lenvatinib resistance in hepatocellular carcinoma by suppressing ACSL1-mediated ferroptosis. BMC Cancer 2025;25:1638.
- 62. Prawira A, Le TBU, Ho RZW, Huynh H. Upregulation of the ErbB family by EZH2 in hepatocellular carcinoma confers resistance to FGFR inhibitor. J Cancer Res Clin Oncol 2021;147:2955-2968.
- 63. Yang Y, Zhang Y, Cao J, et al. FGFR4 and EZH2 inhibitors synergistically induce hepatocellular carcinoma apoptosis via repressing YAP signaling. J Exp Clin Cancer Res 2023;42:96.
- 64. Zhou J, Che J, Xu L, et al. Enhancer of zeste homolog 2 promotes hepatocellular cancer progression and chemoresistance by enhancing protein kinase B activation through microRNA-381-mediated SET domain bifurcated 1. Bioengineered 2022;13:5737-5755.
- 65. Zhang Q, Deng X, Tang X, et al. MicroRNA-20a suppresses tumor proliferation and metastasis in hepatocellular carcinoma by directly targeting EZH1. Front Oncol 2021;11:737986.
- 66. Zeng T, Luo L, Huang Y, Ye X, Lin J. Upregulation of miR-138 increases sensitivity to cisplatin in hepatocellular carcinoma by regulating EZH2. Biomed Res Int 2021;2021:6665918.
- 67. Chen J, Xu Z, Huang H, Tang Y, Shan H, Xiao F. SETD1A drives stemness by reprogramming the epigenetic landscape in hepatocellular carcinoma stem cells. JCI Insight 2023;8:e168375.
- 68. Wang C, Xia Z, Li Z, et al. Expression of SET domain bifurcated histone lysine methyltransferase 1 and its clinical prognostic significance in hepatocellular carcinoma. J Clin Lab Anal 2022;36:e24090.
- 69. Kato H, Hayami S, Ueno M, et al. Histone methyltransferase SUV420H1/KMT5B contributes to poor prognosis in hepatocellular carcinoma. Cancer Sci 2024;115:385-400.
- 70. Wu SY, Xie ZY, Yan LY, et al. The correlation of EZH2 expression with the progression and prognosis of hepatocellular carcinoma. BMC Immunol 2022;23:28.
- 71. Chen SY, Li JD, Huang ZG, et al. Clinical significance of upregulation of EZH1 expression in hepatocellular carcinoma tissues. J Gastrointestin Liver Dis 2024;33:44-56.
- 72. Zhang Y, Lao W, Yang K, et al. SUV39H1 is a novel biomarker targeting oxidative phosphorylation in hepatitis B virus-associated hepatocellular carcinoma. BMC Cancer 2023;23:1159.
- 73. Jin Y, Rao K, Zheng J, Zhang X, Luo Y, He J. Deficiency of TET2-mediated KMT2D self-transcription confers a targetable vulnerability in hepatocellular carcinoma. PNAS Nexus 2024;3:pgae504.
- 74. Tsuchiya J, Miyoshi M, Kakinuma S, et al. Hepatitis B Virus-KMT2B integration drives hepatic oncogenic processes in a human gene-edited induced pluripotent stem cells-derived model. Cell Mol Gastroenterol Hepatol 2025;19:101422.
- 75. Wang X, He M, Zhou L, Chen W. EZH2 expression in hepatocellular carcinoma and its relationship with circadian rhythm-related genes. Sci Rep 2025;15:42177.
- 76. Bai B, Liu Y, Fu XM, et al. Dysregulation of EZH2/miR-138-5p axis contributes to radiosensitivity in hepatocellular carcinoma cell by downregulating hypoxia-inducible factor 1 alpha (HIF-1alpha). Oxid Med Cell Longev 2022;2022:7608712.